+86-28-82633987sales@biopurify.com


SakuranetinCAS No.:2957-21-3
|
||||||||||
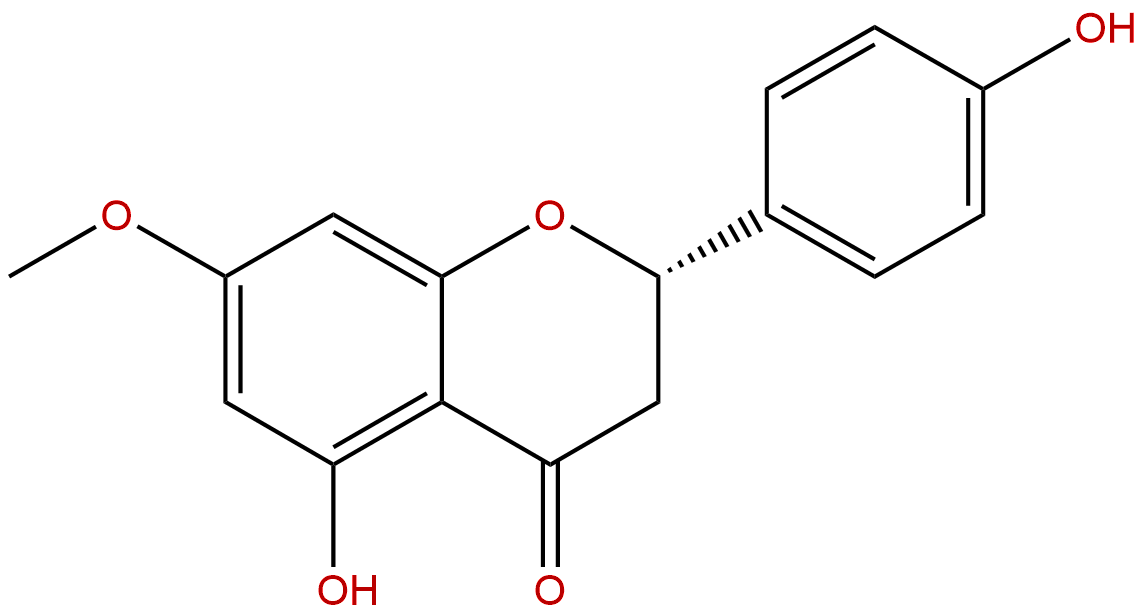 |
|
|
||||||||
| Catalogue No.: | BP1243 |
| Formula: | C16H14O5 |
| Mol Weight: | 286.283 |
Synonym name:
Catalogue No.: BP1243
Cas No.: 2957-21-3
Formula: C16H14O5
Mol Weight: 286.283
Botanical Source: Widespread in the Compositae (Baccharis, Eupatorium, Gymnopentzia, Polymnia), also in Iris, Betula, Notholaena, Prunus, Eucalyptus, Juglans, Poecilanthe and other genera
Purity: 95%~99%
Analysis Method: HPLC-DAD or/and HPLC-ELSD
Identification Method: Mass, NMR
Packing: Brown vial or HDPE plastic bottle
Can be supplied from milligrams to grams.
For Reference Standard and R&D, Not for Human Use Directly.
Inquire for bulk scale.
Description:
Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves, it has antifungal, antimutagenic, anti-inflammatory and antioxidant effects, it shows a suppressive effect on umu gene expression of SOS response in Salmonella typhimurium TA1535/pSK1002 against the mutagen furylfuramide. Sakuranetin may contribute to maintain glucose homeostasis in animals, it can strongly induce differentiation of 3T3-L1 preadipocytes, stimulate glucose uptake in differentiated 3T3-L1 adipocytes.Sakuranetin stimulates melanogenesis in B16 melanoma cells via inhibition of ERK1/2 and PI3K/AKT signaling pathways, which lead to upregulation of Tyr, TRP1, and TRP2.
References:
Molecules. 2014 Aug 4;19(8):11404-18.
Analysis on blast fungus-responsive characters of a flavonoid phytoalexin sakuranetin; accumulation in infected rice leaves, antifungal activity and detoxification by fungus.
To understand the role of the rice flavonoid phytoalexin (PA) Sakuranetin for blast resistance, the fungus-responsive characteristics were studied.
METHODS AND RESULTS:
Young rice leaves in a resistant line exhibited hypersensitive reaction (HR) within 3 days post inoculation (dpi) of a spore suspension, and an increase in Sakuranetin was detected at 3 dpi, increasing to 4-fold at 4 dpi. In the susceptible line, increased Sakuranetin was detected at 4 dpi, but not at 3 dpi, by which a large fungus mass has accumulated without HR. Induced expression of a PA biosynthesis gene OsNOMT for naringenin 7-O-methyltransferase was found before accumulation of Sakuranetin in both cultivars. The antifungal activity of Sakuranetin was considerably higher than that of the major rice diterpenoid PA momilactone A in vitro and in vivo under similar experimental conditions. The decrease and detoxification of Sakuranetin were detected in both solid and liquid mycelium cultures, and they took place slower than those of momilactone A. Estimated local concentration of Sakuranetin at HR lesions was thought to be effective for fungus restriction, while that at enlarged lesions in susceptible rice was insufficient.
CONCLUSIONS:
These results indicate possible involvement of Sakuranetin in blast resistance and its specific relation to blast fungus.
Biochem Biophys Res Commun. 2008 Aug 8;372(4):835-9.
Sakuranetin induces adipogenesis of 3T3-L1 cells through enhanced expression of PPARgamma2.
Sakuranetin (5,4'-dihydroxy-7-methoxyflavone) belongs to the flavanone class of polyphenols predominantly known as phytoalexin in rice plant.
METHODS AND RESULTS:
In this study, we demonstrate that Sakuranetin strongly induces differentiation of 3T3-L1 preadipocytes, as evidenced by increased triglyceride accumulation and glycerol-3-phosphate dehydrogenase (GPDH) activity. In addition, even in the absence of adipogenic hormonal stimuli, Sakuranetin strongly induced adipogenesis and expression of genes that are critical for the adipocytes phenotype. Time-course analyses indicated that Sakuranetin induces PPARgamma2 expression without prior induction of C/EBPbeta, a transcriptional regulator of PPARgamma2 in adipogenesis. In 3T3-L1 preadipocytes, the transcriptional factors GATA-2 and GATA-3 are known to down-regulate adipogenesis by direct binding to the C/EBPbeta protein and to the GATA-binding site on the PPARgamma2 promoter. We found that Sakuranetin significantly reduced the expression of GATA-2. Moreover, we observed that Sakuranetin stimulated glucose uptake in differentiated 3T3-L1 adipocytes.
CONCLUSIONS:
These results suggest that Sakuranetin may contribute to maintain glucose homeostasis in animals.
J. Food Sci., 2003, 68(68):52-6.
Antimutagenic Activity of Sakuranetin from Prunus Jamasakura.
Sakuranetin (compound 1) from bark of Prunus jamasakura showed a suppressive effect on umu gene expression of SOS response in Salmonella typhimurium TA1535/pSK1002 against the mutagen furylfuramide.
METHODS AND RESULTS:
Gene expression was suppressed 83% at a concentration of 0.70 μmol/mL. The ID50 value of compound 1 was 0.30 μmol/mL. This compound showed the suppression of 4NQO, MNNG, Trp-P-1, AfB1, activated Trp-P-1, and UV irradiation-induced SOS response. The methylated derivative (compound 2) of compound 1 showed less suppressive effect against all mutagens than compound 1. The antimutagenic activities of compounds 1 and 2 against furylfuramide, Trp-P-1, and activated Trp-P-1 were assayed by the Ames test using the S. typhimurium TA100 strain.
Acta Histochem. 2016 Jul;118(6):615-624.
Sakuranetin reverses vascular peribronchial and lung parenchyma remodeling in a murine model of chronic allergic pulmonary inflammation.
Asthma is a disease of high prevalence and morbidity that generates high costs in hospitalization and treatment. Although the airway is involved in the physiopathology of asthma, there is also evidence of the importance of vascular and lung parenchyma inflammation and remodeling, which can contribute to the functional pulmonary alterations observed in asthmatic patients. Our aim was to evaluate treatment using Sakuranetin, a flavone isolated from the twigs of Baccharis retusa (Asteraceae), on vascular and lung parenchyma alterations in an experimental murine model of asthma.
METHODS AND RESULTS:
Male BALB/c mice were subjected to a sensitization protocol with ovalbumin for 30days and were treated with or without Sakuranetin (20mg/kg/mice) or dexamethasone (5mg/kg/mice); then, the lungs were collected for histopathological analysis. We evaluated extracellular matrix remodeling (collagen and elastic fibers), inflammation (eosinophils and NF-kB) and oxidative stress (8-isoprostane) in the pulmonary vessels and lung parenchyma. The thickness of the vascular wall was quantified, as well as the vascular endothelial growth factor (VEGF) levels. We demonstrated that Sakuranetin reduced the number of eosinophils and elastic fibers in both the pulmonary vessels and the lung parenchyma, probably due to a reduction of oxidative stress and of the transcription factor NF-kB and VEGF levels in the lung. In addition, it reduced the thickness of the pulmonary vascular wall. The treatment had no effect on the collagen fibers. In most of the parameters, the effect of Sakuranetin was similar to the dexamethasone effect.
CONCLUSIONS:
Sakuranetin had anti-inflammatory and antioxidant effects, preventing vascular and distal parenchyma changes in this experimental model of asthma.
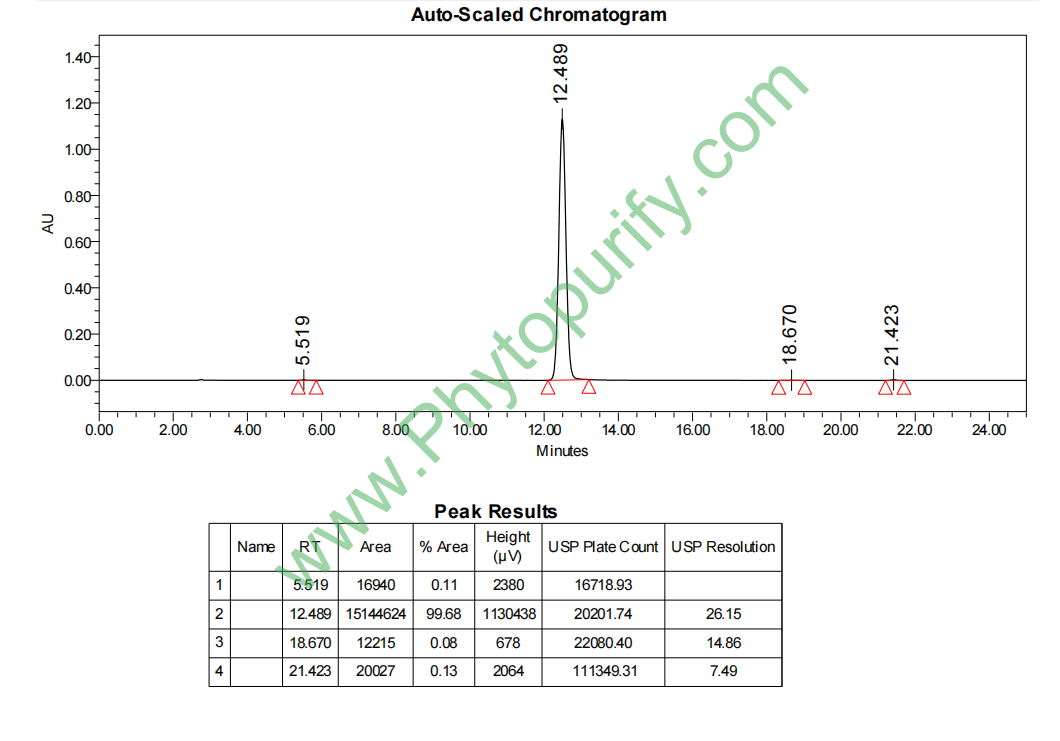
HPLC of Sakuranetin
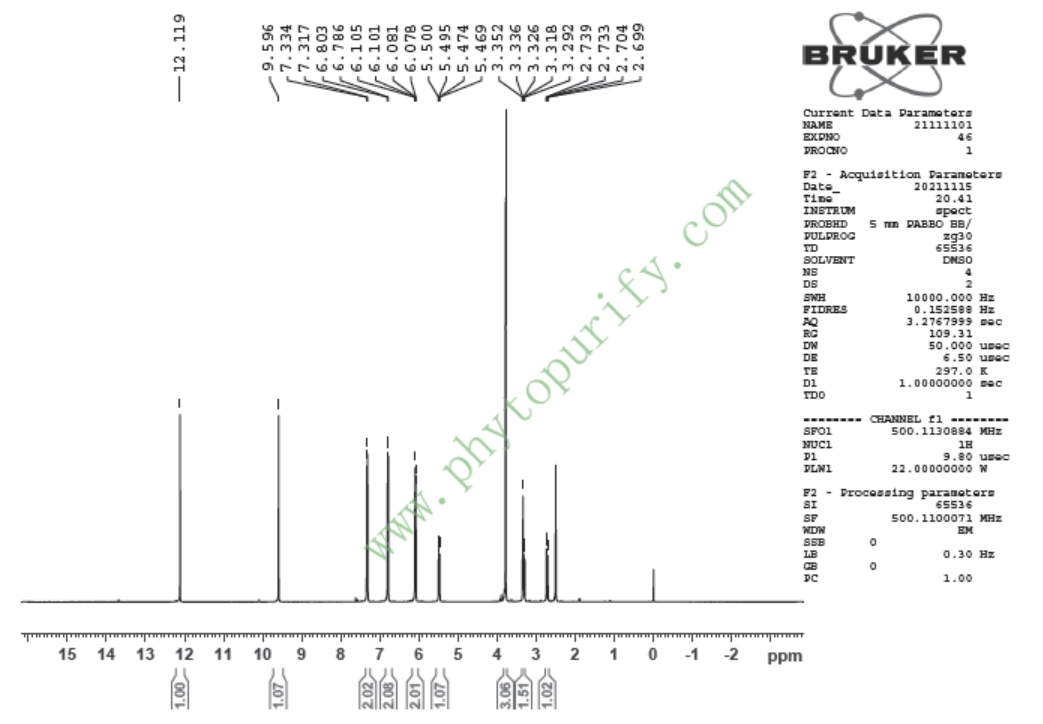
HNMR of Sakuranetin
 |






